
L’effetto nocivo delle temperature elevate (ipertermia) sulle piante è strettamente legato con altri fattori ambientali come la luce, l’umidità atmosferica e la disponibilità di acqua nel suolo. Si tratta di un’interdipendenza molto stretta ed è molto difficoltoso distinguere gli effetti dei singoli fattori sulla pianta. E’ evidente, ad esempio, che l’innalzamento della temperatura potrà provocare risposte delle foglie sensibilmente differenti in funzione dell’umidità atmosferica. Sarà inoltre determinante la possibilità della pianta di poter compensare l’aumento di traspirazione con l’assorbimento radicale, correlando quindi gli effetti dello stress termico con il contenuto di umidità nel suolo.
I danni dello stress termico
I danni da ipertermia sui vegetali possono essere distinti in:
- Danni secondari: quando dipendono sostanzialmente dal manifestarsi dello stress idrico;
- Danni primari indiretti: si manifestano in genere in seguito ad una esposizione continua e a lungo prolungata di temperature moderatamente elevate;
- Danni primari diretti: sono quelli che avvengono in conseguenza di shock termici di breve durata, a temperature più elevate.
L’aumento delle temperature, oltre i limiti fisiologici delle piante, può rallentare o bloccare la crescita dei vegetali con conseguenti danni alla produttività, senza che apparente si manifestino sintomi visibili. In questa fase il ripristino di condizioni ambientali più favorevoli possono ancora limitare i danni economici alla coltura.
L’innalzamento termico può anche arrivare a causare un blocco irreversibile nella crescita qualora giunga a provocare danni metabolici irreversibili. Il processo fotosintetico, in particolare, è particolarmente sensibile alle alte temperature, potendosi bloccare a livelli termici parecchio più bassi di quelli che provocano l’arresto del processo respiratorio. Di conseguenza anche quando i valori termici provocano l’arresto della fotosintesi, il processo respiratorio può comunque continuare con consumo delle riserve sino all’esaurimento.
Sono stati osservati anche effetti rilevanti dello stress termico sulla quantità e la qualità delle proteine. Le temperature elevate possono interferire con il metabolismo proteico giungendo, in casi estremi, alla denaturazione delle stesse o alla coagulazione. Effetti negativi si possono avere anche sui lipidi che compongono le membrane delle cellule e di altre strutture cellulari come i mitocondri e i cloroplasti, organuli che sono rispettivamente sede dei processi respiratori e fotosintetici.
La capacità delle piante di rispondere all’innalzamento termico con meccanismi di termoregolazione è piuttosto limitata e la regolazione della temperatura è sostanzialmente dovuto all’effetto raffreddante della traspirazione. La temperatura delle piante dipende pertanto essenzialmente dall’ambiente nelle sue due componenti: aria e terreno. Si può qui citare, a titolo di esempio, uno studio condotto da Edgington e Walker (1957) sulle fluttuazioni giornaliere della temperatura interna di piante di pomodoro in relazione all’intensità di traspirazione. Si è visto che l’aumento della traspirazione, massima nelle ore diurne, coincide con una diminuzione della temperatura interna della pianta per effetto del raffreddamento esercitato dal flusso idrico alla temperatura del terreno. Si sono osservate differenze di temperatura di 10°C e più tra le temperature interne di fusti di piante vive e morte in piena insolazione.
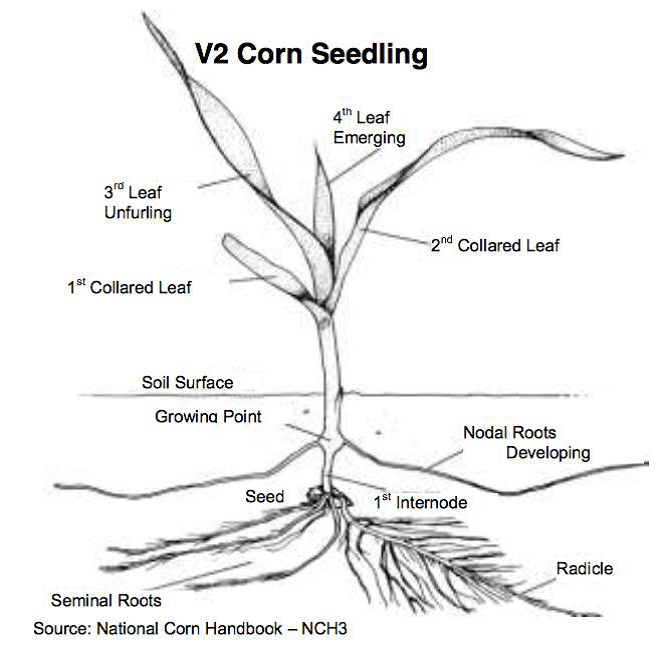
Il mais
Nel mais la temperatura ottimale è compresa tra i 24° ed i 30°C in funzione della disponibilità idrica e dello stadio vegetativo della pianta. I valori massimi sono richiesti in corrispondenza della fioritura – allegagione (Giardini A., 1989).
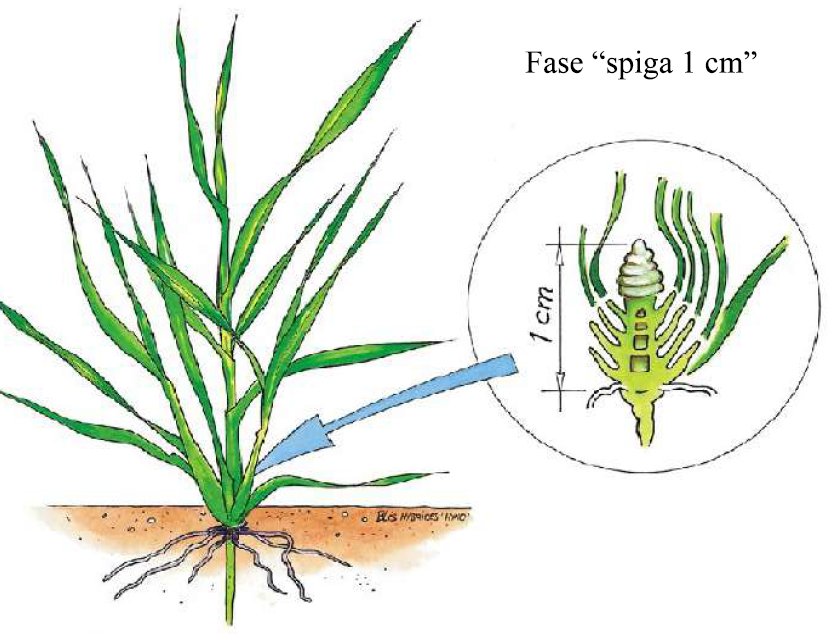
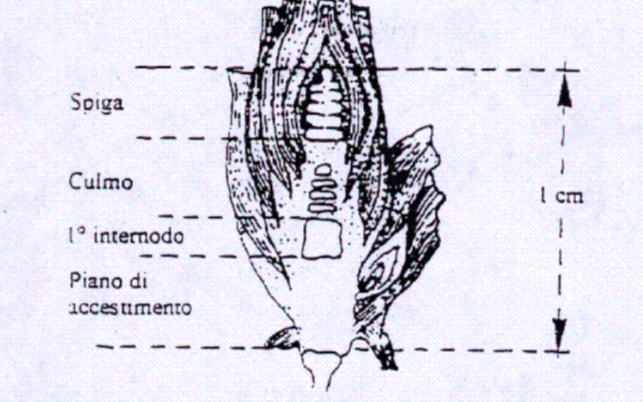
Fioritura – impollinazione
La fase di fioritura – impollinazione è quella in cui lo stress termico può maggiormente compromettere le future rese della coltura. Le alte temperature spesso si accompagnano allo stress idrico, tanto che non è sempre facile separarne i singoli effetti. Si può comunque affermare che qualora nel terreno sia presente una adeguata umidità, le alte temperature di per sé non mettano a dura prova l’impollinazione.
La fioritura femminile
Le sete che emergono dalla spiga sono, come è noto, gli stigmi dei fiori femminili. Ogni seta è collegata a un singolo ovulo, cioè ad una potenziale cariosside. Le sete iniziano a svilupparsi 10-14 giorni prima che si rendano visibili emergendo dalle brattee che avvolgono la spiga. L’allungamento delle sete inizia da prima dagli ovuli basali delle spiga, quindi procede progressivamente verso l’apice.

Una volta emerse continuano ad accrescersi piuttosto velocemente per 1-2 giorni, per poi rallentare progressivamente nei giorni successivi. Comunque il loro allungamento si interrompe poco dopo che un granulo di polline si deposita su di esse germinando. Se non impollinate, l’accrescimento delle sete si interromperebbe una decina di giorni successivamente all’emergenza; sete insolitamente lunghe possono essere, infatti, un indicatore di una cattiva impollinazione della spiga.

Lo sviluppo delle sete solitamente avviene in sincronia con il rilascio del polline, agevolando così l’impollinazione. Le condizioni di stress possono interferire con questa sincronia. La siccità in questa delicata fase è il più pericoloso fattore di stress, soprattutto quando è accompagnata da alte temperature e bassa umidità atmosferica. Le sete hanno un elevato contenuto in acqua, maggiore di un qualsiasi altro tessuto vegetale. Sono quindi molto sensibili alle variazioni di umidità nella pianta e quindi alla siccità. Questa può ostacolare o rallentare il loro allungamento ritardandone la fuoriuscita oppure, nelle peggiori condizioni ambientali, impedendola. Si avrà quindi una cattiva impollinazione delle spighe qualora il rilascio del polline è ormai finito o è insufficiente.
Nelle condizioni di stress termico e idrico con bassa umidità atmosferica le sete possono facilmente essiccare, perdendo la recettività al polline.
Alcuni ibridi selezionati per la tolleranza alla siccità, avendo una crescita delle sete più veloce in condizioni di stress idrico, consentono di prevenire questo problema. Tuttavia non mancano le “controindicazioni”. In condizioni ottimali di sviluppo delle sete, questa maggiore rapidità di sviluppo può anticipare di 4-5 giorni la loro comparsa, anticipando sensibilmente il rilascio del polline, riproponendo i problemi di mancata allegagione.
La fioritura maschile
Anche il rilascio del polline (antesi) e la sua vitalità, quindi la sua capacità fecondante, possono essere seriamente compromessi dalle elevate temperature unite alla siccità.
Il polline viene rilasciato per lo più dalla prima mattinata fino a metà di essa, quando la temperatura è ancora relativamente bassa. Un secondo rilascio può talvolta avvenire nel tardo pomeriggio con temperature più fresche. Umidità e temperature sono due fattori chiave durante la deiscenza. E’ stata osservata una riduzione della dispersione del polline quando le temperature superano i 30°C. I granuli di polline rimangono vitali per 18 – 24 ore in condizioni favorevoli, mentre la loro vitalità si riduce a un paio d’ore in condizioni di caldo estremo. Temperature superiori a 37°C causano un forte stress al polline che può disseccare ancor prima di fecondare l’ovulo. Le alte temperature possono anche ridurre la vitalità del polline prima che venga rilasciato dalle antere.
Accrescimento delle cariossidi
Abbiamo ricordato più sopra come le elevate temperature diurne possano causare una riduzione della capacità fotosintetica. Questo aspetto, presente anche nel mais, determina una riduzione della produzione di zuccheri destinata a ripercuotersi sulla produzione finale del mais. Qualora le temperature elevate persistano durante la notte il danno produttivo si aggrava. In queste condizioni nelle ore notturne l’attività respiratoria si mantiene elevata con conseguente consumo di zuccheri. Temperature più basse di notte in genere favoriscono anche la traslocazione dei carboidrati verso gli organi di riserva (cariossidi).

Successivamente alla fecondazione, condizioni di siccità o stress termico possono provocare l’aborto degli ovuli fecondati. I chicchi abortiti hanno piccole dimensioni, spesso di colore biancastro. Le cariossidi sono maggiormente suscettibili a questo danno nelle due settimane successive alla fecondazione, in particolare lo sono quelle prossime alla punta della spiga. Va comunque ricordato che l’aborto delle cariossidi può anche essere provocato da qualsiasi fenomeno che limiti l’attività fotosintetica, quindi la disponibilità di fotosintetati, durante o subito dopo la fecondazione. Oltre gli stress da calore e da siccità, possono essere chiamati in causa, quali responsabili del danno, le carenze nutrizionali (soprattutto azoto), le perdite del tessuto fogliare dovute a patogeni o a grandine ecc.

Stress idrici e termici possono portare ad una prematura morte della pianta con precoce arresto dello sviluppo delle cariossidi.
Le perdite di resa, approssimativamente calcolate, conseguenti alla prematura morte delle foglie, ma non degli steli, si stimano essere:
- 36%, quando la morte si verifica allo stadio R4 (maturazione cerosa),
- 31%, allo stadio R5 (formazione del dente nella cariosside),
- 7%, a metà della linea del latte dello sviluppo della cariosside.
Bibliografia
Goidànich G. (1983): Manuale di patologia vegetale, volume primo. Edizioni Agricole Bologna.
Matta A., Pennazio S. (1984) Elementi di fisiopatologia vegetale, Pitagora editrice Bologna.
Baldoni R., Giardini L. (1989): Coltivazioni erbacee. Patron Editore Bologna.
Fonti web consultate il 23-07-2022 :
http://www.kingcorn.org/news/timeless/Silks.html http://www.kingcorn.org/news/timeless/GrainFillStress.html








































 Adulto P. japonica
Adulto P. japonica

















